![An Australian Government Initiative [logo]](/images/austgovt_brown_90px.gif)

|

Form and structure
This page will be devoted to explaining the common lichen structures you can see with the naked eye as well as giving some basic information about the internal structure of lichens. To help illustrate some of the points there will be links to drawings taken from Albert Schneider's A text-book of general lichenology, published in 1897, and there's more about those drawings (and some warnings) on the SCHNEIDER'S BOOK page.
When you look at a lichen such as the foliose Xanthoria filsonii ![]() you see a yellowish basal sheet on which there are numerous darker yellow disks. The basal sheet is the thallus (composed of fungal and photobiont tissue) and the disks are the apothecia (in which the fungal partner produces sexual spores). In some genera the apothecia are narrow and elongated (sometimes even branching) and are then termed lirellae
you see a yellowish basal sheet on which there are numerous darker yellow disks. The basal sheet is the thallus (composed of fungal and photobiont tissue) and the disks are the apothecia (in which the fungal partner produces sexual spores). In some genera the apothecia are narrow and elongated (sometimes even branching) and are then termed lirellae ![]()
![]() . Another type of fungal fruiting body is a perithecium which, to the naked eye, looks like a small, hemispherical pimple, typically black
. Another type of fungal fruiting body is a perithecium which, to the naked eye, looks like a small, hemispherical pimple, typically black ![]() . Within a perithecium are the asci in which sexual spores are produced. The surface of a thallus may show minute, powdery granules (called soredia), each soredium consisting of a few photobiont cells surrounded by fungal filaments. Also, the thallus may produce tiny, simple or branched spiny outgrowths (called isidia), again a mixture of fungal and photobiont cells. The isidia are easily broken and both they and the soredia are easily dispersed and contain everything needed to produce new thalli. Soredia and isidia are means of vegetative reproduction. There is a separate REPRODUCTIVE STRUCTURES page and the rest of the current page will concentrate on the non-reproductive thallus features. Furthermore, in almost all lichens the mycobiont is an ascomycete and this page will be devoted to the forms and structures in the ascolichens. There is a separate page about BASIDIOLICHENS which contains details of both vegetative and reproductive structures of such lichens
. Within a perithecium are the asci in which sexual spores are produced. The surface of a thallus may show minute, powdery granules (called soredia), each soredium consisting of a few photobiont cells surrounded by fungal filaments. Also, the thallus may produce tiny, simple or branched spiny outgrowths (called isidia), again a mixture of fungal and photobiont cells. The isidia are easily broken and both they and the soredia are easily dispersed and contain everything needed to produce new thalli. Soredia and isidia are means of vegetative reproduction. There is a separate REPRODUCTIVE STRUCTURES page and the rest of the current page will concentrate on the non-reproductive thallus features. Furthermore, in almost all lichens the mycobiont is an ascomycete and this page will be devoted to the forms and structures in the ascolichens. There is a separate page about BASIDIOLICHENS which contains details of both vegetative and reproductive structures of such lichens![]() .
.
The WHAT IS A LICHEN page introduced the three major thallus growth forms: crustose, foliose and fruticose. In the first two the thallus has an upper (or dorsal) surface and a lower (or ventral) surface whereas in the third the terms upper and lower are often meaningless when talking about the thallus. Typically there is just an outside surface to a fruticose thallus. In a number of genera the thallus has components of two different forms and such a thallus is called a dimorphic thallus. Cladonia is an example of a very common genus in which the species have dimorphic thalli and there will be some more about Cladonia a little later on this page.
Thallus structure
The following very simplistic cross-sectional diagrams show the broad internal compositions of many of the crustose, foliose and fruticose lichens. Leftmost is a crustose species. Black indicates the cortex, a band of compacted fungal hyphae that serves as a protective skin. Below the cortex, in green, is a layer of photobiont cells. If you carefully scrape away the upper cortex you can see the photobiont layer - green if it's algal and blue-green if its cyanobacterial. This photo ![]() shows several thalli of Endocarpon pusillum, with some of the upper cortex of one scraped away. Below the photobiont layer is the medulla, a loose weave of fungal hyphae, and shown here in grey. You can also see some grey hyphae growing down from the medulla and these hyphae penetrate the substrate and so anchor the thallus very tightly. The middle diagram shows a foliose lichen. An immediate difference from the crustose lichen diagram is that the foliose thallus shown here has two cortices - an upper cortex and a lower cortex beneath the medulla. The foliose diagram has an additional colour. The blue lines indicate rhizines, which are bundles of hyphae. These penetrate the substrate and anchor the thallus. Fruticose lichens are typically highly branched with the branches often more or less circular in cross section. The final diagram shows a cross section of the branch of a fruticose lichen. There is a single cortex which constitutes the outermost band, within which there is a concentric photobiont band and the medulla occupies the central portion of the branch.
shows several thalli of Endocarpon pusillum, with some of the upper cortex of one scraped away. Below the photobiont layer is the medulla, a loose weave of fungal hyphae, and shown here in grey. You can also see some grey hyphae growing down from the medulla and these hyphae penetrate the substrate and so anchor the thallus very tightly. The middle diagram shows a foliose lichen. An immediate difference from the crustose lichen diagram is that the foliose thallus shown here has two cortices - an upper cortex and a lower cortex beneath the medulla. The foliose diagram has an additional colour. The blue lines indicate rhizines, which are bundles of hyphae. These penetrate the substrate and anchor the thallus. Fruticose lichens are typically highly branched with the branches often more or less circular in cross section. The final diagram shows a cross section of the branch of a fruticose lichen. There is a single cortex which constitutes the outermost band, within which there is a concentric photobiont band and the medulla occupies the central portion of the branch.
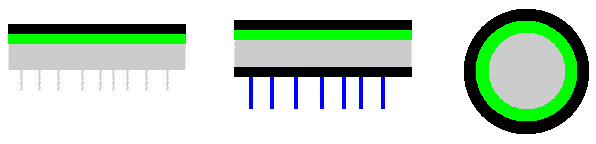
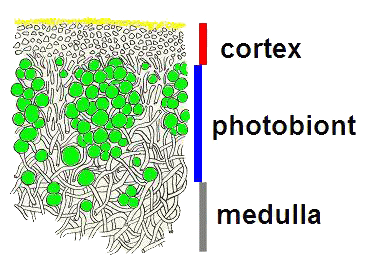
Before going on I stress again that these diagrams are very simplistic and serve only to convey the relative positions of cortices, medullae, photobionts and rhizines in the thalli of many lichens. It is worth noting that in a number of species cortices may bear crystalline deposits, derived from secondary metabolites produced by the lichens. Shortly I will go into a little more detail which will let you see some of the variation in thallus structure - as well as exceptions to some of the above statements. Before doing so, I'll give a more realistic diagram of part of a lichen thallus after the end of this paragraph. In this species the cortex is composed of two layers, a thin upper layer of tiny, yellow granules and a thicker, colourless layer below that. Below the cortex are the spherical cells of the photobiont (shown here in green) and below that the medulla. In the medulla the separate, filamentous hyphae are easy to see but in the lower layer of the cortex the hyphae are compacted![]() .
.
Though a crustose thallus has distinct upper and lower surfaces, the tight binding to the substrate results in the lower surface not being visible. Rhizines on a foliose thallus may be dense or sparse and there are foliose species without rhizines. A lichen with rhizines is termed rhizinate and an erhizinate one lacks rhizines. When rhizines are present (and regardless of whether they are dense or sparse) they may be found anywhere under the thallus in some species while in other species the rhizines are confined to certain areas. This variation in rhizine density and placement means that some foliose thalli are fairly loosely attached to their substrates while others are very firmly attached. In some foliose species the thallus is attached only centrally ![]() , leaving the rest of the thallus free of the substrate and so allowing the underside to be seen easily. Other foliose species have thalli so well attached over much of their under-surface that they might initially be mistaken for crustose species. There is also variety in rhizine shape between species. Rhizines may be just simple linear bundles as shown in the diagram above, However they may also be anything from sparsely to richly branched. In some species the rhizines fork and may do so repeatedly. In others each rhizine has a main axis but with short lateral branches coming off the main axis and yet others resemble well-worn paint brushes that have bristles going off in all directions. As already mentioned some foliose species have no rhizines and Xanthoparmelia semiviridis is a striking example. The thalli sit loosely on the soil, curled up when dry
, leaving the rest of the thallus free of the substrate and so allowing the underside to be seen easily. Other foliose species have thalli so well attached over much of their under-surface that they might initially be mistaken for crustose species. There is also variety in rhizine shape between species. Rhizines may be just simple linear bundles as shown in the diagram above, However they may also be anything from sparsely to richly branched. In some species the rhizines fork and may do so repeatedly. In others each rhizine has a main axis but with short lateral branches coming off the main axis and yet others resemble well-worn paint brushes that have bristles going off in all directions. As already mentioned some foliose species have no rhizines and Xanthoparmelia semiviridis is a striking example. The thalli sit loosely on the soil, curled up when dry ![]() but flat and with a darker colour when moist
but flat and with a darker colour when moist ![]() .
.
In a number of species hyphae may grow out from a cortex to give the cortex a hairy appearance. Amongst the hairy species there are those in which the hairs are sparse and at the other extreme those in which the hairs are dense. There is also variation in the structure and arrangement of such hairs and there are technical descriptive terms for all these variations, but there is no point in going into that sort of detail on this website.
Cortices vary considerably in their structure and there are cortex-less lichens, such as various byssoid lichens. A byssoid thallus ![]() has a somewhat wispy appearance, like cotton-wool teased out to some degree, and is composed of loosely interwoven hyphae and photobiont cells. When present, cortices vary from thin to relatively thick depending on the species. Schneider provided drawings of FLIMSY thalli as well as of a VARIETY of more robust thalli. The first diagram of a foliose thallus on this page showed it as having two cortices and that is common, but there are foliose lichens that lack a lower cortex. An Australian example is Heterodea muelleri
has a somewhat wispy appearance, like cotton-wool teased out to some degree, and is composed of loosely interwoven hyphae and photobiont cells. When present, cortices vary from thin to relatively thick depending on the species. Schneider provided drawings of FLIMSY thalli as well as of a VARIETY of more robust thalli. The first diagram of a foliose thallus on this page showed it as having two cortices and that is common, but there are foliose lichens that lack a lower cortex. An Australian example is Heterodea muelleri ![]() , a species that is widespread and fairly common in southern Australia. Though it lacks a lower cortex, the thallus does possess rhizines.
, a species that is widespread and fairly common in southern Australia. Though it lacks a lower cortex, the thallus does possess rhizines.
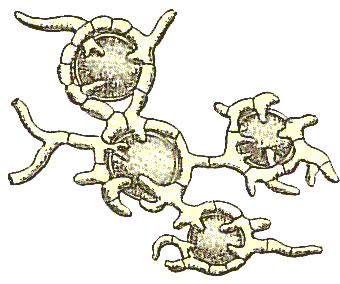
In each of the simple diagrams above, the photobiont cells are shown as being confined to a well-defined band immediately below a cortex and distinct from the medulla. That is the case in a great many lichens (and such lichens are described as heteromerous) but there are a few genera in which the photobiont cells are distributed randomly through the thallus and Schneider included drawings of a such species, described as homoiomerous. Whatever the arrangement, fungal hyphae 'harvest' a large proportion of the nutrients produced by the photobiont and the following illustrations (again, taken from Schneider's book) show fungal hyphae branching and encircling a number of globose photobiont cells. There is some variation in the way the hyphae attach themselves to or penetrate the photobiont cells in order to harvest the photosynthetic products.
Podetia and pseudopodetia
 Cladonia is a very widespread and common lichen genus in which many species develop initially as squamulose thalli. A squamule is a small, flake-like thallus and a colony of a squamulose lichen looks like a scattering of small flakes or scales on the substrate. However, the species of Cladonia also produce upright fruticose structures called podetia - sometimes with the appearance of fairly simple stalks, sometimes flared at the apex and so presenting a somewhat trumpet-like form. In this photo
Cladonia is a very widespread and common lichen genus in which many species develop initially as squamulose thalli. A squamule is a small, flake-like thallus and a colony of a squamulose lichen looks like a scattering of small flakes or scales on the substrate. However, the species of Cladonia also produce upright fruticose structures called podetia - sometimes with the appearance of fairly simple stalks, sometimes flared at the apex and so presenting a somewhat trumpet-like form. In this photo ![]() you can see a Cladonia colony growing on soil. There are numerous squamules on the soil but you can also see a number of the upright podetia with broader apices. Moreover, at the margins of some of the broader apices you can see additional podetia developing and there are also squamulose growths on the podetia as shown in this enlargement
you can see a Cladonia colony growing on soil. There are numerous squamules on the soil but you can also see a number of the upright podetia with broader apices. Moreover, at the margins of some of the broader apices you can see additional podetia developing and there are also squamulose growths on the podetia as shown in this enlargement ![]() of part of the previous photo. On the right is a considerable enlargement of another part of the first photo. On the right is a considerable enlargement of part of the previous photo. The yellow arrows point to the lobes of a squamulose thallus from which a podetium is growing. This young podetium is a few millimetres tall. The white arrow points to what looks to be another thallus that it is in contact with the one indicated by the yellow arrows, rather than part of the yellow-arrow thallus. Squamules, such as the ones on the soil around the bases of podetia are often referred to as basal squamules. In a number of Cladonia species the basal squamules are evanescent and won't be seen once the podetia have developed. Here
of part of the previous photo. On the right is a considerable enlargement of another part of the first photo. On the right is a considerable enlargement of part of the previous photo. The yellow arrows point to the lobes of a squamulose thallus from which a podetium is growing. This young podetium is a few millimetres tall. The white arrow points to what looks to be another thallus that it is in contact with the one indicated by the yellow arrows, rather than part of the yellow-arrow thallus. Squamules, such as the ones on the soil around the bases of podetia are often referred to as basal squamules. In a number of Cladonia species the basal squamules are evanescent and won't be seen once the podetia have developed. Here ![]() and here
and here ![]() are other examples of Cladonia podetia. The coloured areas atop the podetia are apothecia. The hyphae that help form the podetia are formed from generative tissue. In a lichen this tissue surrounds the sexual organs and gives rise to the fungal spore-producing structures (such as apothecia). Thus, in developmental terms the whole podetium can be viewed as part of the fruiting body. There are a number of lichen genera in which apothecia are held aloft on stalks but where the stalk is composed of the vegetative tissue of the thallus and where the generative tissue is found only at the top of the stalk. In such a case the stalk is simply an elongation of the thallus and, though it may be superficially podetium-like, is developmentally quite different and is called a pseudopodetium. For example, the brown apothecia of Stereocaulon ramulosum
are other examples of Cladonia podetia. The coloured areas atop the podetia are apothecia. The hyphae that help form the podetia are formed from generative tissue. In a lichen this tissue surrounds the sexual organs and gives rise to the fungal spore-producing structures (such as apothecia). Thus, in developmental terms the whole podetium can be viewed as part of the fruiting body. There are a number of lichen genera in which apothecia are held aloft on stalks but where the stalk is composed of the vegetative tissue of the thallus and where the generative tissue is found only at the top of the stalk. In such a case the stalk is simply an elongation of the thallus and, though it may be superficially podetium-like, is developmentally quite different and is called a pseudopodetium. For example, the brown apothecia of Stereocaulon ramulosum ![]() are produced on pseudopodetia. Many Stereocaulon species have dimorphic thalli, with a non-persistent primary thallus (often crustose or squamulose) from which the fruticose, and often highly branched, pseudopodetia develop. In the genus Cladia the thalli are composed of perforate to reticulate pseudopodetia
are produced on pseudopodetia. Many Stereocaulon species have dimorphic thalli, with a non-persistent primary thallus (often crustose or squamulose) from which the fruticose, and often highly branched, pseudopodetia develop. In the genus Cladia the thalli are composed of perforate to reticulate pseudopodetia ![]() , so exposing the medulla to contact with the outside air
, so exposing the medulla to contact with the outside air![]() .
.
Cyphellae, pseudocyphellae and maculae
Cyphellae and pseudocyphellae are breaks in the cortex of a thallus but are of two different forms. To the naked eye a cyphella looks like a smooth-surfaced depression in the thallus. The surface of the depression is lined with fungal cells different in form to the hyphae of the medulla. While a pseudocyphella is also a break in a cortex it is one where, in contrast to a cyphella, the hyphae of the medulla are exposed to the air, rather than having a differentiated hyphal layer separating the medulla from the outside. You can find Schneider's diagrams of cyphella and pseudocyphella structures HERE. Cyphellae are found only in the foliose genus Sticta ![]() and are almost always confined to the underside of the thallus. Pseudocyphellae are found in a small number of genera of foliose, fruticose and even crustose lichens. When present in foliose lichens pseudocyphellae
and are almost always confined to the underside of the thallus. Pseudocyphellae are found in a small number of genera of foliose, fruticose and even crustose lichens. When present in foliose lichens pseudocyphellae ![]() may be found on lower or upper surfaces depending on the species. Cyphellae and pseudocyphellae are gas exchange sites.
may be found on lower or upper surfaces depending on the species. Cyphellae and pseudocyphellae are gas exchange sites.
In lichens with a photobiont layer beneath a cortex, the photobiont layer need not be continuous. Since the photobiont layer does contribute to the overall colour of the thallus, an area free of photobiont will show as a paler spot or line on the thallus. Such spots or lines, usually irregular in shape, are called maculae (singular: macula).
Mixed photobionts
A number of lichens have an alga as the major photobiont but with a cyanobacterium present as a minor photobiont in discrete pockets and such a pocket is called a cephalodium. Cephalodia may be found within the thallus or as growths on the thallus ![]() . In a number of lichens free-living cyanobacterial cells can become trapped in hairy cortices and additional hyphae then grow around such cells to eventually develop external cephalodia. Internal cephalodia are common in the genera Lobaria and Sticta where they develop as lumps in the medulla. These cephalodia can become so large as to cause visible swellings of the thallus surface. External cephalodia vary in shape between species and may be disk-like, rounded or even branched. In some species the delimited areas are less delimited than in others, so leading to minor differences of opinion. For example, the northern hemisphere species Solorina crocea has both algal and cyanobacterial photobionts, with the latter found in a patchy layer below the algal layer. I have seen one book describe the species as having a double layer of photobionts, whereas another book describes the patches as cephalodia. In the species Compsocladium archboldianum (known from Irian Jaya and Papua New Guinea) algal cells form the photobiont layer below the cortex and individual cyanobacterial filaments are found in the medulla.
. In a number of lichens free-living cyanobacterial cells can become trapped in hairy cortices and additional hyphae then grow around such cells to eventually develop external cephalodia. Internal cephalodia are common in the genera Lobaria and Sticta where they develop as lumps in the medulla. These cephalodia can become so large as to cause visible swellings of the thallus surface. External cephalodia vary in shape between species and may be disk-like, rounded or even branched. In some species the delimited areas are less delimited than in others, so leading to minor differences of opinion. For example, the northern hemisphere species Solorina crocea has both algal and cyanobacterial photobionts, with the latter found in a patchy layer below the algal layer. I have seen one book describe the species as having a double layer of photobionts, whereas another book describes the patches as cephalodia. In the species Compsocladium archboldianum (known from Irian Jaya and Papua New Guinea) algal cells form the photobiont layer below the cortex and individual cyanobacterial filaments are found in the medulla.
Note that some references define a cephalodium as a delimited region of one type of photobiont in a lichen where the major photobiont is of the other type, so allowing for algal pockets in a lichen with the dominant photobiont a cyanobacterium. However, no such example appears to be known, but even if such examples exist the most common arrangement would still be for a cephalodium to consist of a cyanobacterial photobiont in a lichen where an alga is the dominant photobiont.
What's dominant - fungus or photobiont?
In the majority of lichens the fungal tissue constitutes the bulk of the thallus, as suggested by the simple diagrams near the top of this page. There is a small number of genera in which photobiont tissue dominates. A number of byssoid genera, with cotton-wool-like thalli fall into this category as do some more robust genera such as Collema and Leptogium. In the latter two the photobiont is cyanobacterial in the genus Nostoc and the thalli of Collema and Leptogium could initially be mistaken for Nostoc, especially if the lichens lacked apothecia. The following photos show Nostoc (left) and a fertile Collema (right). Without the white apothecia the Collema thallus is featureless and gelatinous in texture, just like a Nostoc.
 |
 |
Given that fungal tissue is usually dominant it would be understandable to conclude that, in such cases, it is the fungus that determines whether a thallus will be crustose, foliose or fruticose. However, the case of STICTA AND 'DENDRISCOCAULON' is instructive. It shows one fungus associating with different photobionts (one an alga, the other a cyanobacterium), but without the cyanobacterium confined to cephalodia. Instead there is a dimorphic thallus, with one form of thallus containing the alga and the other form the cyanobacterium, so that thallus form is influenced by the photobiont. Note that in Cladonia and Stereocaulon the different components of the dimorphic thallus have the same photobiont, though you will also find cephalodia in many species of Stereocaulon.
Form and structure pages on this website Crustose lichens |