
Rein Orchids
Deciduous terrestrials with fleshy tubers and an erect stem on which flat or channelled smooth leaves are arranged in a rosette at the base or near the centre of the stem, sometimes just 1 or 2 at the base. The unbranched inflorescence carries white, green or yellowish stalked flowers in a terminal raceme. The dorsal sepal overlaps with the petals (which can be lobed) to form a hood over the column. The labellum, which has a basal spur, is usually 3-lobed with the lobes short or sometimes long and thread-like. The column has 2 stigmas with elongate stalked lobes that are not fused to the labellum base.
Similar Genera
Significant Generic Characters
Deciduous autotrophic (rarely mycotrophic) terrestrial (rarely epiphytic) orchids; tubers fleshy, subterranean, bearing rhizoids; leaves few-many, flat or conduplicate, smooth, sheathing at the base, non-articulate, spiral or distichous, variably arranged on the stem; inflorescence racemose, terminal; flowers resupinate, mostly white or green; dorsal sepal overlapping with the petals to form a galea; petals entire, lobed or bifid; labellum usually 3-lobed, with a nectariferous basal spur; pollinaria 2, each with a sectile pollinium on a caudicle and a naked viscidium; stigmata 2.
Size and Distribution
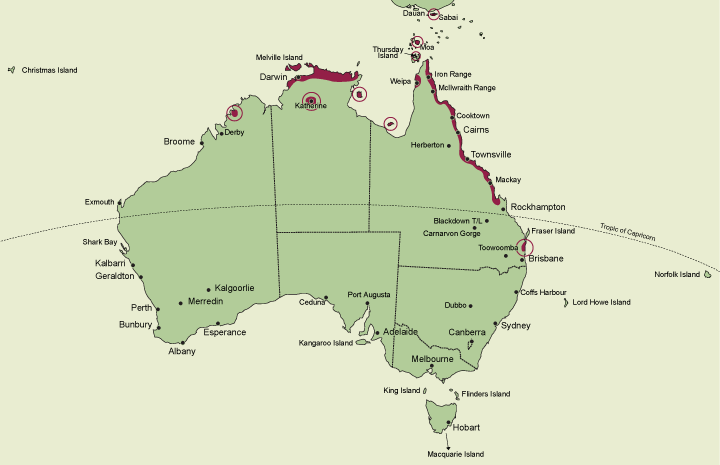
As currently interpreted, Habenaria is a large unwieldy genus of 600-10000 species distributed in tropical and subtropical regions of the world. Seventeen species occur in Australia, all but one restricted to northern tropical areas. Habenaria harroldii is the exception occurring in the subtropics of southeastern Queensland. State occurrence: Queensland (including Moa, Saibai and Thursday Islands), Northern Territory, Western Australia.
Ecology
The native species of Habenaria grow in a range of habitats including rainforest, seasonal wetland dominated by Melaleuca and Pandanus, swamp forest, grassland, heathland, coastal forest and on coastal headlands. Whereas the majority of species grow in low-lying areas that become wet to inundated during the summer monsoon, a number grow in well-drained soils.
Biology
Pollination: The flowers of most native species of Habenaria are insect-pollinated but the vectors are unknown. The flowers are mostly white, greenish or yellowish and are often fragrant, especially in the late afternoon and early evening and the floral spurs contain nectar at the distal end. A number of exotic species of Habenaria are pollinated by moths or mosquitoes which insert their mouthparts into the labellum spur to feed on the nectar, the pollinia in some cases becoming attached to the insect�s eye.Similar syndromes may apply to some of the native species.
Reproduction: Reproduction in the native species of Habenaria is mainly from seed although most, if not all, produce daughter tubers and can form small vegetative groups or loose colonies. Seed dispersal takes 4-8 weeks from pollination and the capsules develop in an erect position. Apomixis possibly occurs in 2 native species, Habenaria harroldii and Habenaria xanthantha.
Seasonal Growth: The majority of native species of Habenaria, with the exception of Habenaria harroldii, occur in the tropics. The seasonal growth of all tropical species is closely tied in with the wet season. The tubers sprout with the onset of the wet and the plants grow and flower during the wet season and die down with the onset of the dry. Habenaria harroldii behaves similarly.
Flowering: The Australian species of Habenaria flower between December and March.
Hybrids: Natural hybrids are unknown between the native species of Habenaria.
Fire: Some native species of Habenaria grow in fire-prone habitats, others grow in sites that are not burnt. While there is no direct stimulation by fire it has been observed that flowering appears to be improved in burnt habitat, especially overgrown habitats that have not been burnt for a number of years.
Derivation
The name Habenaria is derived from the Latin habena, reins, and refers to the long filamentous divisions on the petals and labellum of some species.
Botanical Description
Perennial geophytic herbs (rarely epiphytic), autotrophic (rarely mycotrophic), deciduous, sympodial. Plants glabrous. Flowering and non-flowering plants monomorphic. Tubers fleshy, with numerous rhizoids, subterranean, paired, held vertically or horizontally; replacement tuber formed on a short dropper; daughter tubers absent or produced on horizontal, fleshy, stolonoid roots. Roots filamentous, short to long, fleshy, unbranched. Stem erect, emergent, unbranched, fleshy, without cataphylls. Trichomes absent. Leaves 1-few per shoot, spirally arranged or distichous, scattered with the upper ones reduced in size and bract-like, sometimes forming a basal rosette or central rosette on the scape, sometimes 1 or 2 basal leaves, flat or conduplicate, smooth, non-articulate, entire, sessile; base sheathing; lower leaves often reduced to imbricate, tubular sheathing bracts. Venation unknown. Inflorescence a terminal raceme, few-many-flowered. Peduncle similar to or longer than the rhachis. Rhachis similar to or shorter than the peduncle, straight. Floral bracts small to large, often foliaceous. Pedicel short, merging with the ovary. Ovary straight, not twisted, narrow. Flowers resupinate, often crowded, small to relatively large, white, green or yellow, often fragrant, lasting a few days, opening sequentially in a spiral. Perianth segments narrow to relatively broad. Dorsal sepal free, dissimilar to the lateral sepals, erect, generally broad and cucullate, concave, overlapping with the petals to form a galea. Lateral sepals free, dissimilar to the dorsal sepal, porrect, deflexed or divergent. Petals free, overlapping with the dorsal sepal to form a galea, entire, bilobed or bifid. Labellum fixed to the base of the column, markedly dissimilar in size and shape to the sepals and petals, calcarate. Labellum lamina 3-lobed, often deeply so (sometimes pelorial and unlobed); lateral lobes small to large, sometimes lobed, narrow to filiform, porrect to widely divergent; midlobe narrow to broad. Spur short to long, subglobose to cylindrical, straight or curved, often distally dilated. Callus obscure, vestigial or absent. Nectar present. Column porrect from the end of the ovary, very short, lacking free filament and style. Column wings absent or present as lateral papillate auricles or appendages. Column foot absent. Pseudospur absent. Anther dorsal, erect or reclinate, fused basally with the column, 2-celled, erostrate, the cells extended at the base into thecal tubes which enclose the caudicles and are adnate to the rostellum lobes. Pollinaria 2. Pollinia 2, narrow, sectile, attached by long narrow caudicles at right angles to the pollinia. Viscidium usually 2, naked, separate or sometimes both fused together, terminal, more or less hemispherical, sometimes enclosed in a bursicle. Rostellum entire or usually lobed, situated between the thecal tubes and the stigmatophores. Stigmata convex, entire or 2-lobed, the lobes (stigmaphores) short or long, free, generally elongate, cylindrical or clavate, hollow. Capsules dehiscent, glabrous, erect; peduncle not elongated in fruit; pedicel not elongated in fruit. Seeds numerous, light coloured, winged.
Taxonomy
Habenaria is included in the subtribe Habenariinae. As currently defined the genus is polyphyletic and in need of a complete study based on molecular and morphological characters (see Bateman et al. 2001). Several natural groups with recogniseable features can be discerned within the genus and at least 4 distinct groups occur in Australia. It is likely from preliminary studies that some or all of these will be recognised at generic rank and it is probable that Habenaria s/s does not occur in Australia.
Notes
The column structure in this genus, particularly details of the anther, stigma, rostellum and pollinarium is complex and the various parts can be difficult to discern and identify.
Nomenclature
Habenaria Willd., Sp. pl. 4(1): 5, 44 (1805). Type species: (Orchis habenaria L.) Habenaria macroceratitis Willd.
Infrageneric Taxa: Many sections have been described in this complex genus but the relationships within and between the groups require complete reassessment. For a summary of the sectional treatment see Cribb (2001).
References
Bateman, R. M., Hollingsworth, P.M., Preston, J., Yi-bo, L., Pridgeon, A.M. and Chase, M.W. (2001). Phylogenetics of Orchideae in Genera Orchidacearum. Volume 2, Orchidoideae (Part 1), 224-232, edited by A.M.Pridgeon, P.C.Cribb and M.W.Chase.
Cribb, P.C. (2001). Habenaria in Genera Orchidacearum. Volume 2, Orchidoideae (Part 1), 298-303, edited by A.M.Pridgeon, P.C.Cribb and M.W.Chase.
Dixon, K. (1994). Tropical terrestrial orchids of Western Australia. Orchadian 11(4): 166-170.
Dockrill, A.W. (1969). Australian Indigemous Orchids. Volume 1. The Society for Growing Australian Plants, Halstead Press, Sydney.
Dockrill, A.W (1992). Australian Indigenous Orchids. Volume 1 & 2. Surrey Beatty & Sons in association with the Society for Growing Australian Plants, Chipping Norton, NSW.
Jones, D.L. (1998). A new species of Habenaria Willd. (Orchidaceae) from southern Queensland. Orchadian 12(9): 414-415.
Jones, D.L. (1998). Two new species of Habenaria Willd. (Orchidaceae) from northern Queensland. Orchadian 12(10): 461-465.
Jones, D.L. (1998). Four new species of Habenaria Willd. (Orchidaceae) from Australia and a new combination for a species from New Guinea. Orchadian 13(11): 516-524.
Lavarack, P.S. (1981). A new species of Habenaria from Cape York Peninsula. Orchadian 7(12): 278-279.
Lavarack, P.S. & Dockrill, A.W. (1999). A new species of Habenaria Willd. (Orchidaceae) from north Queensland. Austrobaileya 5(2): 331-335.